即时发布
北卡罗来纳州立大学和北卡罗来纳大学查普尔山研究者发现两位重要校对蛋白如何知道在脱氧核糖核酸复制时哪里查找错误,以及他们如何协同信号人体修复机制
细胞准备分治时,DNA先分治双螺旋“解片化为2个独立的骨架新核素-adenine、cytosine、guanine或emine-填入骨架另一端的漏洞,并配对对应方(九元甲型和九元甲型)并复制脱氧核素拷贝新老细胞核素大都匹配,但偶发-百万分之一-不匹配
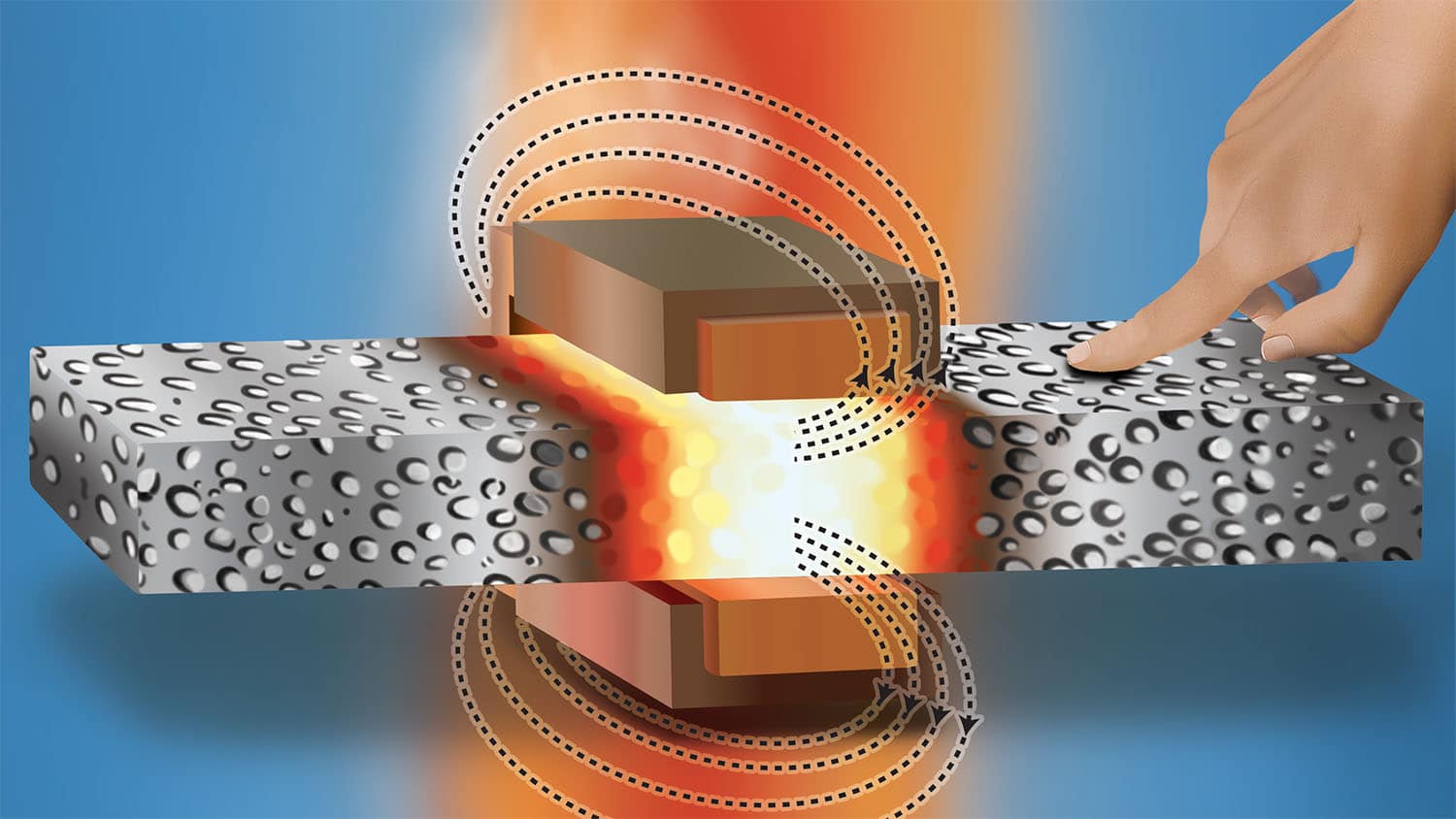
幸运的是,我们的身体有检测修复系统-双蛋白类MutS和MutLmats仿真校对后沿新生成的脱氧核糖核酸链侧滑动发现不匹配时,它锁定出错网站并招聘MutL加入mutl新合成脱氧核糖核酸链中插取nick标志它有缺陷并信号用异质填充含有误差的脱氧核糖核酸核酸匹配重开 填充空白整个过程减少千倍复制错误, 并成为我们身体 最佳防控基因变异 和问题从中产生,像癌症
所有生物都使用此进程复制并修复DNA, 我们知道元件是什么, 但也有一些关键机制不为人所理解.NC State物理教授Keith Weninger举个例子,当修复脱氧核糖核酸时,它只在脱氧核酸新片段发生,而不是老模版蛋白质知道沿线移动的方程 以清除缺陷通信发生在这里, 但对它如何工作有分歧。”
科学家知道一个叫PCNA的蛋白质 在这个通信中起重要作用PCNA站点脱氧核糖核酸分解,该站点物理定位表示哪个部分脱氧核酸新称它为线性偏差讯号 //////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////MutS和MutL与信号交互作用,允许MutL窃取适当位置并显示修复需求科学家有三种模式进行通信:MTS和MTL组合成锁链向PCNA滑动与信号交互作用,标记区域滑动删除或以相同效果折叠脱氧核糖核酸或MutL以某种方式覆盖有缺陷区域,以这种方式信号修复
温宁格与合著者Dorothy Erie从UNC-Chapel Hill开始确定最可能的通信方法使用单分子荧光法使研究者能够看单片脱氧核糖核酸运动时确定当MTS发现误差时,它改变形状方式允许MTL绑定它,并存放在不匹配网站MutL后修改形状为“grab”另一个mutL,等等,涂层线有缺陷部分并窃取需要修复的区域换句话说,Weninger、Erie和他们的同僚发现第三模型正确
维宁格说道 系统对保持基因组稳定至关紧要和进程开端相识, 工作填充中间步骤之一 完全未知 与某些人所想大相径庭
变异MutS和MutL与非常特殊癌症相关联,if we're to explication why缺陷引致疾病, 我们首先必须理解这些蛋白质通常如何工作研究是通向理解的另一步
研究在线显示国家科学院记录.资金由美国癌症学会、国家卫生研究所和国家科学基金会提供。杜克大学的Paul Modrich、Wesleyan大学的Manju Hingorani和后院生和研究生RuoyiQui(NC州)、Elizabeth Sacho(NC州)、Hunt Wilkins(UNC-CH)、SindgongZhang(Duke)和Miho Sakato(Wesleyan)也为工作做出了贡献。
说话-
向编辑注解:摘要如下
MutL陷阱脱氧核糖核酸匹配
DOI:101073/pnas1505655112
作者类KeithWeninger、RuoyiQue、ElizabethSacho和NC州立大学Dorothy ErieHunterWilkinsUNC-Chapel山保尔莫德里希 兴东张 杜克大学Manju Hingorani,Wesleyan大学MihoSakato
发布在线上网国家科学院记录
抽象性 :
脱氧核糖核酸不匹配修复在所有生物中,除表达 mutH外,脱氧核糖核酸不匹配、MutS、MutL和复制过程性因子(et-cramp或PCNA)交互作用激活隐式mtlendonucleNick为下游修复蛋白质提供切入点mtsm和Mutl集合脱氧核糖核酸以允许MutlnucleProkaryotes和eukaryotes的MutS同族体都接受运动夹定状态的整齐修改,移动夹定状态可以远离不匹配状态然而,MutS移动绑定函数未知与MutL交互生成移动MTS-MutL综合体或错位综合体单分子FRET
确定srmus水生MutL捕捉MutS取非绑定式异动蛋白质组件通常装配MutL多于MutS复杂程序提供本地标识符,与e-cram/PCNA交互可辨别父/女身份发现MutL根本改变MutS动作 后匹配检测重构MMR信号进程当前思维对基因组稳定至关重要